Motility in a Test Tube
Ronald Vale
 Ron Vale
Ron Vale
Ron Vale is a Vice President at Howard Hughes Medical Institute & Executive Director of the Janelia Research Campus where he studies molecular motors, the cytoskeleton and signaling. Ron received his Ph.D. in Neuroscience from Stanford University in 1985. His honors include the Lasker Award in Basic Medical Research, the Shaw Prize in Life Sciences, and the Canada Gairdner Award; he was elected to the US National Academy of Sciences and the National Academy of Medicine. Ron believes in the free dissemination of scientific knowledge throughout the world. He is the founder of iBiology, a non-profit organization that produces free videos of talks by leading scientists, and the founder and executive producer of The Explorer's Guide to Biology.
Summary
Cities bustle with the motion of people and commercial goods. Just as goods must be transported from one point to another within a city, molecules also need to be transported within cells to the destinations where they are needed. Accordingly, cells have developed roadways (cytoskeletal filaments), engines (protein motors), and cargo containers (membrane organelles) to physically transport molecules. These biological transport systems are essential for all cells, but they play especially important roles in neurons, which must transport molecules over very long distances. A failure of the biological transport system can lead to diseases of the nervous system as well as other organs. A cell is about a billion times smaller than a city. How can one study such a miniaturized transport system? Powerful microscopes can be used to visualize small subcellular objects, and the videos in this "Key Experiment" provide a peek at the busy highways inside a nerve cell. In addition, this experiment uses biochemistry to re-build a cell's transportation system in a test tube. These simplified methods for studying motility led to the discovery of a new motor protein (kinesin) and became powerful tools for understanding how biological motility works. This "Key Experiment" also explains "assays" and "controls" (core concepts in biology) and reveals how unexpected twists and turns can shape research projects and careers.
Learning Overview
Big Concepts
Cells contain thousands of proteins performing many processes. Biochemists try to reduce the complexity in understanding these processes by studying a biological activity outside of the cell (in vitro) and with a minimal number of components. In this "Key Experiment," the complex process of membrane organelle transport in a nerve cell was rebuilt in vitro from its molecular parts, providing new insights and tools for understanding the basis of biological motility.
Bio-Dictionary Terms Used
Bacteriophage (phage), electron microscope (EM), enzyme, gel electrophoresis, fractionation, light microscope, lipid, membrane, mitochondria, model organisms, mutation, neuron, neuron cell body, nucleus, plasma membrane, protein, protein domain, protein purification, sucrose, stem cell
Terms and Concepts Explained
Actin, adenosine triphosphate (ATP), assay, ATPases, axon, axoplasmic transport, buffer, cytoskeleton, kinesin, in vitro reconstitution, microtubule, motor neuron, motor protein, organelles, ultracentrifuge, variable
Introduction
-
Neurons have long, narrow tubes called axons that connect the cell body to the nerve terminal. Membrane organelles move within the axons to transport goods between the cell body and nerve terminal through a process called axonal transport.
-
At the time of this experiment (1984), the mechanism of axonal transport was not known.
What Events Preceded the Experiment?
-
It is important to pick a good organism to study a biological problem. For studying axonal transport, squid was a good choice because it has a "giant axon" that can be easily dissected out of the squid.
-
In the early 1980s, Allen, Brady, and Lasek observed organelles moving inside of the axon using a video microscope, one of the first occasions in which video cameras and computers were used to record images from a microscope rather than just looking with the naked eye.
-
The cytoskeleton is a network of protein filaments inside of cells. Two main cytoskeletal systems found in all eukaryotic cells are actin filaments and microtubules. Electron microscopy studies revealed that microtubules serve as tracks for long-distance transport of organelles in axons.
Setting up the Experiment
-
To learn more about how axonal transport works, the investigators isolated the components that they thought were needed for transport and then recombined them in a test tube. They then used a microscope to examine the combined mixture to observe if organelles were transported along microtubules. This type of experiment is called in vitro reconstitution (taking a system apart and putting it back together in a test tube).
-
The isolated components were:
A) Microtubules (the tracks for movement) made from a purified protein called tubulin.
B) Adenosine triphosphate (ATP; the chemical energy source).
C) Membrane organelles (the cargo) from the giant axon, which were isolated by the method of sucrose density gradient ultracentrifugation. An ultracentrifuge spins samples at high speeds, allowing biological materials to be separated on the basis of both size and density.
D) Soluble protein fraction (free proteins that are neither associated with organelles nor the cytoskeleton) were also isolated from the giant axon.
Doing the Key Experiment
-
The in vitro reconstitution experiment involved mixing various combinations of the components described above and then looking through a microscope to observe if the isolated organelles would move along the microtubule tracks.
-
The scientists first combined 1) microtubules, 2) membrane organelles, and 3) ATP. They thought that this combination might work as it was expected that the motors are bound to organelles. However, no movement was observed.
-
Then, they thought that a component might be missing and hence added the soluble protein fraction. Now the organelles started moving along microtubules, suggesting that this fraction contained a key ingredient needed for the movement.
-
The key ingredient in the soluble protein fraction proved to be a motor protein itself. When they combined microtubules, the soluble protein fraction, and ATP (but no organelles), surprisingly, they observed microtubules moving over the surface of a glass microscope slide. A follow-up experiment showed that the motor protein stuck onto the glass and the glass-bound motor proteins propelled the microtubules along the surface.
-
Small plastic beads also could bind to the unknown motor protein from the soluble protein fraction. The motor protein would transport the beads along the microtubules in a manner that looked similar to the transport of membrane organelles inside of axons.
-
This experiment demonstrated that axonal transport could be reconstituted from isolated components. It also led to new experimental strategies (called "assays") for studying motor proteins—the movement of microtubules across a glass surface and the movement of plastic beads along stationary microtubules. These new assays, which did not involve membrane organelles (complex objects themselves), provided new and simple ways of measuring the activity of motor proteins.
What Happened Next?
-
The simplified in vitro motility assays provided means to isolate a new motor protein from the axon, which was named "kinesin."
-
In addition to helping in the identification of a new motor protein, these assays provided powerful tools for understanding how motor proteins convert chemical energy from ATP into movement.
Closing Thoughts
-
Serendipity plays a big role in scientific research. The in vitro motility assays of microtubules moving across a glass surface and plastic beads moving along stationary microtubules were unanticipated at the start of the experiment and proved to be the biggest advancement.
Guided Paper
Vale, R.D., Schnapp, B.J., Reese, T.S., and Sheetz, M.P. Organelle, bead and microtubule translocations promoted by soluble factors from the squid giant axon. Cell 1985;40:559–569.
This paper shows that a protein, now called kinesin, is responsible for the complex process of membrane organelle transport within nerve cells, providing new insights and tools for understanding the basis of biological motility.
DownloadIntroduction
Could a single evening in the laboratory change the course of one's life? This is the story of such an evening.
I was a 25-year-old student in a combined PhD/MD training program at Stanford University. At the time of this story, I was wrapping up the research for my PhD degree and, in 3 weeks, was scheduled to stop my research and transition to the final phase of my MD training. I had already completed enough work to obtain my PhD. But I wanted to end my research work with a bang and not a whimper. But I was not getting any results. Were the experiments too difficult? Or was I just clumsy?
As my experiments were not going well, it was tempting to give up. However, my fascination with the research kept me going. Lying in front of me was a great puzzle that I wanted to solve. The clock was ticking; my scientific research would come to halt for at least 2 years after I returned to medical school. It was now or never.
I was studying how neurons transport materials (proteins and membranes) from the location where they are made to the destinations where they are needed. Most biomolecules are made in the neuron cell body, where the nucleus is located. However, some of these molecules have to be shipped to the nerve terminal. Although all cells need to transport materials, the distance between the cell body and nerve terminal can be much greater than the distance across most other cells in the body. A motor neuron, for example, has its cell body in the spinal cord and its nerve terminal could be located at a muscle in your foot, a distance of around 1 m (Figure 1). Extending between the cell body and the terminal is a very long process called the axon; its length is approximately a million times greater than its diameter. The axon transmits electrical impulses that stimulate and cause muscles to contract. In addition, the interior of the axon is a busy highway transporting molecular goods in both directions between the cell body and nerve terminal.
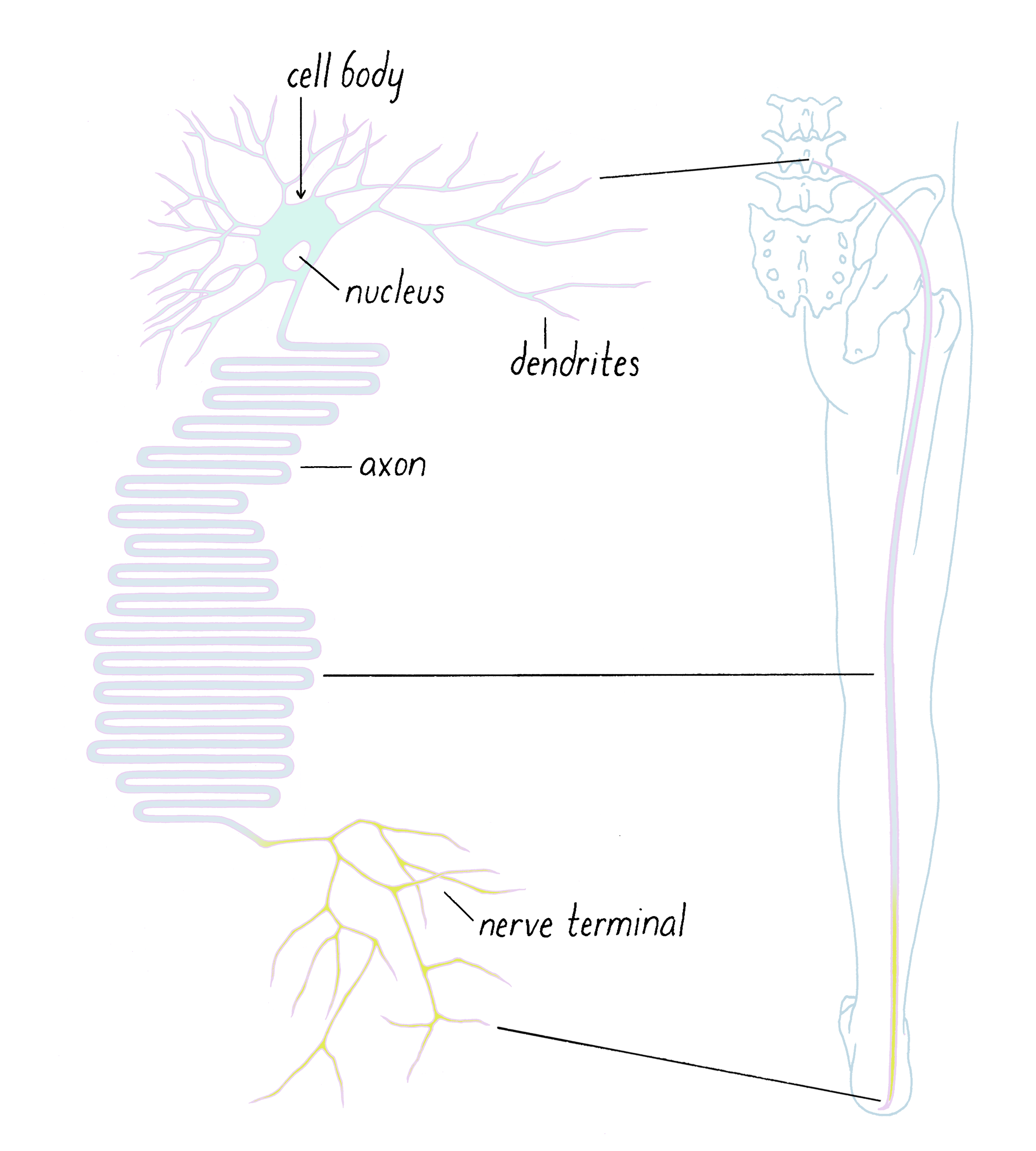
Many of the cellular "goods" are packed in small membrane organelles, which are also referred to as membrane vesicles (Figure 2). These lipid containers act as delivery vans; they move molecules that are attached to their surface or stored in their interior from one place in a cell to another.

Using sophisticated microscopes, scientists could see a tremendous flow of organelles moving in both directions inside of axons (Video 1). If you see a truck moving, you know that there is some sort of engine powering it, and that it is traveling on a road. We suspected that there must be the molecular equivalents of roads and motors that power axonal transport as well. However, in the early 1980s, little was known about what they actually were. That was the puzzle that my colleagues (Mike Sheetz, Bruce Schnapp, and Tom Reese) and I wanted to solve.
I was trying to take apart the axonal transport machinery and then recombine it in a test tube and have it work again. This is called "in vitro reconstitution." If one could control and study the system in this way, one might be able to understand how it works. Imagine taking apart an engine and putting it back together; if you succeed in re-assembling a functional engine, you would certainly understand how an engine works.
Explorer's Question: What features of transport do you notice in this intact axon?
Answer: You might notice that movement occurs in both directions. However, individual small organelles tend to move unidirectionally. The larger, elongated mitochondria tend to move slower, in a halting manner, and exhibit bidirectional motion. There are some faint hints of "tracks" that will be discussed later. You may observe other interesting things in this movie as well.
Now back to my special evening in the laboratory. Four months had passed with essentially no results, and on this day, in particular, I had spent 16 hours in the lab and felt like going home. But I thought that I should squeeze in a quick "control" experiment before calling it a night. Controls are ways of checking the validity of an experiment. However, the control experiment in this story led to something much more than a routine check—it produced a discovery that led me to postpone my return to medical school (indefinitely) and determined the next two decades of my scientific research.
What is a control experiment? And why would someone decide to "drop out" of medical school because of one? Let's find out.
What events preceded the experiment?
To tackle a scientific problem, it is important to choose the right experimental organism. In the case of axonal transport, human neurons might be the most relevant one for understanding human disease; however, people do not want to part with their neurons for scientific research. Today, one can study human neurons derived from human-derived stem cells, but those types of cells were not available at the time.
Even if human neurons were available, the squid was still a better choice as a model organism for studying axonal transport (Video 2). The reason is that squid possesses a "giant axon," which, at ~0.5 mm, is >100-fold larger in diameter than most human axons. The giant axon enables the squid to propel itself rapidly away from its predators by contracting its body muscles. The giant axon is big enough to be seen by the naked eye and dissected out of the squid.
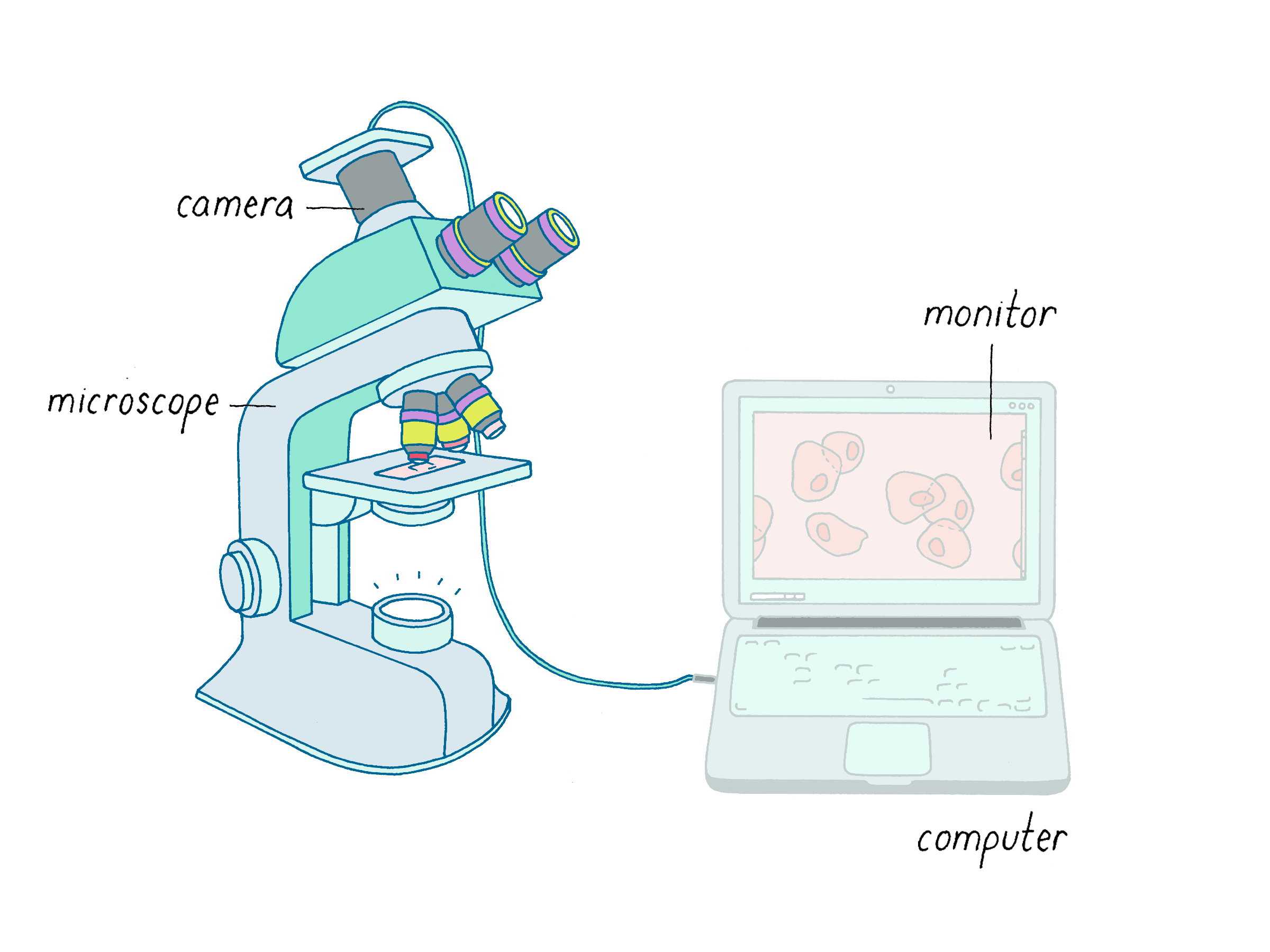
If you want to use squid for scientific research, then you have to travel to where they can be caught (squids do not travel well by car or plane). The unofficial squid capital of the United States is the beautiful Marine Biological Laboratory (MBL) in the small town of Woods Hole, Massachusetts. Between May and October, an MBL fishing boat collects and delivers squid to a sea water tank in your laboratory. In 1982, Robert Allen, Scott Brady, and Ray Lasek put squid to great use. Using Allen's newly developed analog video microscope (Figure 3), these researchers recorded amazing movies of transport occurring inside of the giant axon (see Video 1). Video microscopy was brand new, cutting-edge technology in an era well before digital cameras and YouTube. I had never seen anything like it when I was a graduate student at Stanford. After seeing these movies, I have been in love with microscopy ever since. As the American baseball player and comedic philosopher Yogi Berra said, "You can observe a lot by just watching."
The organelles in an intact axon (Video 1) were moving mostly along straight paths. Hence, scientists suspected that they might be transported along some type of cellular track. When our group, as well as Robert Allen's, removed the material inside of the axon (axoplasm), dissociated it (broke it apart), and looked under the microscope, we could clearly see individual tracks and membrane organelles moving along them (Video 3).
Explorer's Question: What features of transport do you observe when the axon was dissociated in a test tube?
Answer: In this movie, most organelles move in one direction along a filament. However, there are various cases of an organelle moving in the opposite direction and they can pass one another without colliding suggesting that the filaments are multi-lane highways. Some organelles (particularly the mitochondria) can attach to two filaments at the same time, indicating they have more than one attachment point. Once bound, membrane organelles can move a long way before detaching. There is also "bouncy" motion off the filament. We will discuss this motion more in Video 9. You may observe other interesting things in this movie as well.
What were these tracks? The best candidates were filaments of the cytoskeleton (Video 4). There are two major kinds of cytoskeletal filaments called actin filaments and microtubules (Figure 4). These filaments are composed of thousands of copies of a protein (actin for actin filaments and tubulin for microtubules) interacting in a head-to-tail manner (Video 4).
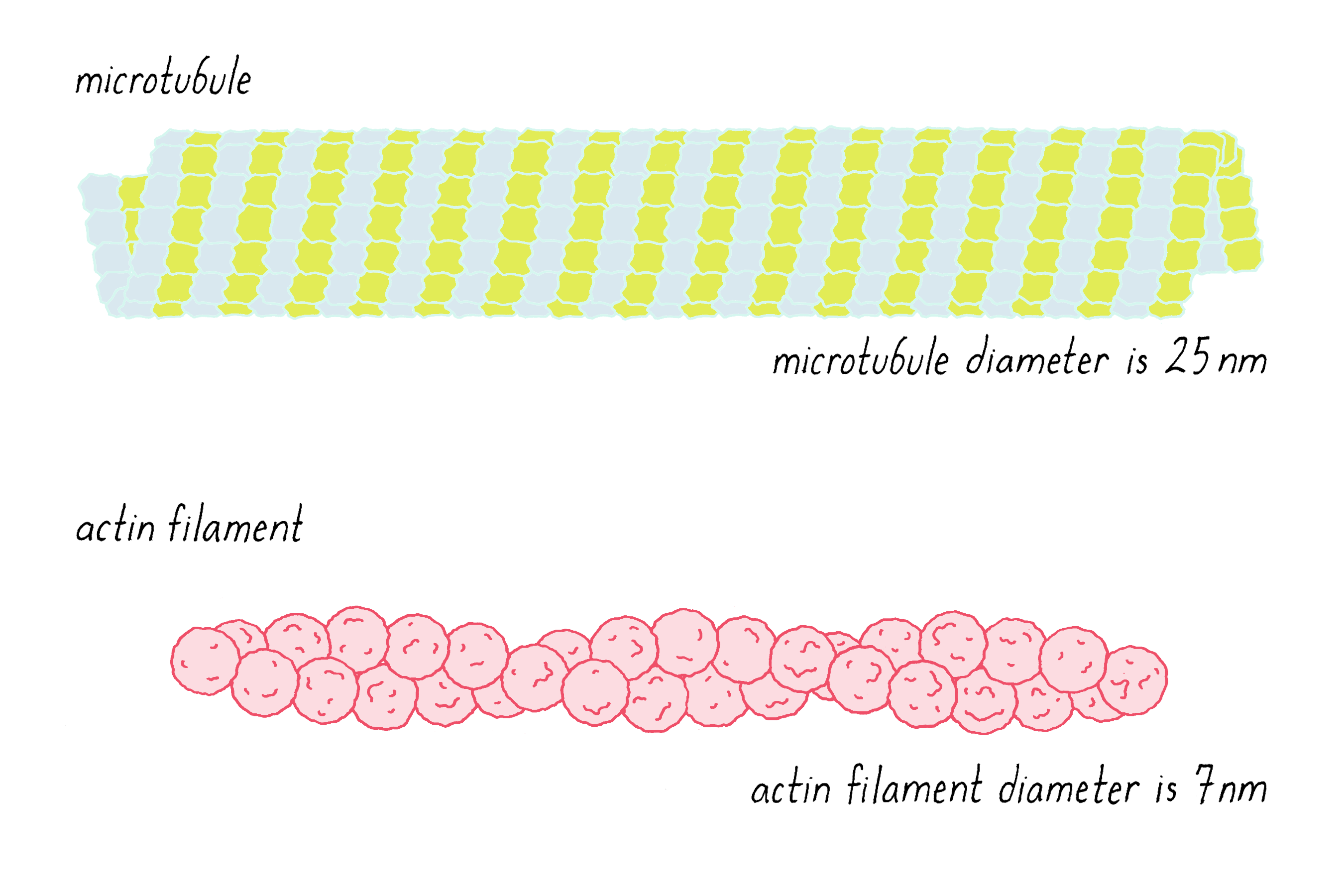
The light microscope, which was used to make the images in Videos 1 and 3, lacks the resolution to discern the details of the tracks shown in Figure 4. Thus, we could not be certain whether they were actin, microtubules, or a combination of the two. We had to turn to a higher resolution technique called electron microscopy (EM). Using EM, Bruce Schnapp and Tom Reese found that the tracks had the diameter and features characteristic of microtubules.
We now knew that organelles were moving along microtubules. But there had to be an additional component—a protein that converts chemical energy into unidirectional motion along the track. Such a protein is referred to as a motor protein. But there are many thousands of different kinds of proteins in a cell—how were we going to identify which protein was the motor protein responsible for transporting organelles?
Setting up the experiment
The quest to identify the motor protein(s) was not easy. How does one identify and study something that one cannot see? In the summer of 1984, I worked on identifying the motor protein through an "in vitro reconstitution" approach.
An in vitro reconstitution involves breaking a cell down into smaller components, and putting various components back together in a test tube ("in vitro") to see if they can perform the desired activity. This would point us to the component that contained our motor protein(s). We use the term "test tube" somewhat figuratively. Most of the in vitro work described below refers to mixing components first in a tube but the experimental mixture was then removed from the tube, applied as a liquid drop on a microscope slide, and then examined under a microscope.
To perform an in vitro reconstitution, it is important to have good inventory of the parts that you will need. Here is what we knew about the parts list for axonal transport in 1984 (see also Video 5, which describes how these components were prepared).
1) Microtubules (the tracks for movement):
As described earlier, we knew that microtubules (Figure 4) were tracks for organelle transport. Microtubules from the squid axon, however, are coated with additional proteins called microtubule-associated proteins that might complicate our experiments. To simplify matters, we wanted to see if organelles could move along microtubules without any microtubule-associated proteins. Using techniques of protein purification, we purified tubulin and reassembled them into microtubules (see Dig Deeper 1). Removing microtubule-associated proteins was a gamble. If they were required for motility, then reconstituting organelle transport then would have failed without them. However, if organelle transport worked without them, then one would have a more simplified system for studying transport. One of the goals of a reconstitution experiment is to simplify a complex cell and identify a minimal parts list required for a biological process.
2) Adenosine Triphosphate (ATP, the source of chemical energy):
ATP (Figure 5) is a small organic molecule that provides the energy required for many life-sustaining processes. It is sometimes referred to as the energy currency of cells. Certain enzymes, called ATPases, hydrolyze the bond linking the second and third phosphates; the energy released by breaking this bond can be coupled to other energy-requiring processes (in our case, to the energy-requiring process of moving along a microtubule).

We needed to supply ATP to power the organelle movement in our reconstitution reaction. Conveniently, ATP could be purchased from a chemical supply company.
3) Isolated membrane organelles:
Small vesicular organelles constitute the majority of cargos moving inside of the axon, although there are also mitochondria (the more elongated objects in Video 2) and some other organelles. See Video 6 to learn more about the organelles inside of a cell. We suspected that the organelles would have protein motors bound to their surface, which would move them along the microtubule.
For our "test tube" reconstitution experiment, we needed to isolate the membrane organelles from the squid giant axon to add them to our test tube reaction. The organelle preparation began by dissecting ten or more giant axons from squid (see Video 5). We then separated the inside of the squid axon (the axoplasm, which we wanted) from the outside plasma membrane (which we threw away). To accomplish this, one end of the axon was cut and the inside was squeezed out, very much like squeezing out toothpaste from a tube (shown in Video 5). Remarkably, the isolated axoplasm still behaves like a gel due to its thick network of cytoskeletal filaments. One can pick up the axoplasm with forceps (Video 5), demonstrating that the inside of cells is not a watery soup, as people sometimes think.
We then collected the axoplasm in a tube with a solution referred to as a buffer solution or a buffer. The buffer is the "special sauce" of biochemistry, a collection of dissolved chemicals designed to keep biochemical reactions working in a test tube. If you want to learn more about buffer solutions, see Dig Deeper 2.
Next, we released the small organelles from the thick "gel" of the axoplasm. We did so by passing the axoplasm vigorously through the narrow opening of a pipette tip. This shears and breaks apart the network of cytoskeletal filaments and makes the axoplasm less gel-like. To move the axoplasm into and out of the pipette tip, we used a pipetteman (Figure 6), a device for dispensing small volumes of liquids (e.g., 1 µL to 1 mL). The dispersed axoplasm is called the axoplasmic homogenate.

Next, the membrane organelles were separated from other contents of the axoplasm. The contents of a cell are very complex, but can be separated into different fractions based on size and density using an ultracentrifuge, a machine that spins samples at >100,000 revolutions per min. At these speeds, the ultracentrifuge generates centripetal forces that can be 100,000 times greater than that of earth's gravity (see Video 5 for a demonstration of the centrifuge used in the experiment). The bigger and denser the biological object, the greater the force it will experience and the faster it will move toward the bottom of the centrifuge tube.
By centrifuging the axoplasm through solutions of different densities, we could collect the membrane organelles at a defined location in the centrifuge tube (Figure 7). We first layered the centrifuge tube with solutions containing different percentages of sucrose (35% sucrose means 35 grams of sucrose dissolved in 100 mL of a solution). The addition of sucrose increases the density of the solution. Membrane organelles are denser than 15% sucrose but lighter than 35% sucrose. During centrifugation, the membrane organelles travel through the 15% sucrose layer, but they do not move into the 35% sucrose layer. At this boundary, a buoyancy force counteracts the centrifugal force and causes the membrane organelles to "float" at the top of the 35% sucrose layer. If you tried to push a cork (low-density object) under water (a higher density solution), you have experienced a buoyancy force. After finishing the centrifugation run, I collected the membrane organelle fraction, which looked like a murky band, on top of the 35% sucrose layer.
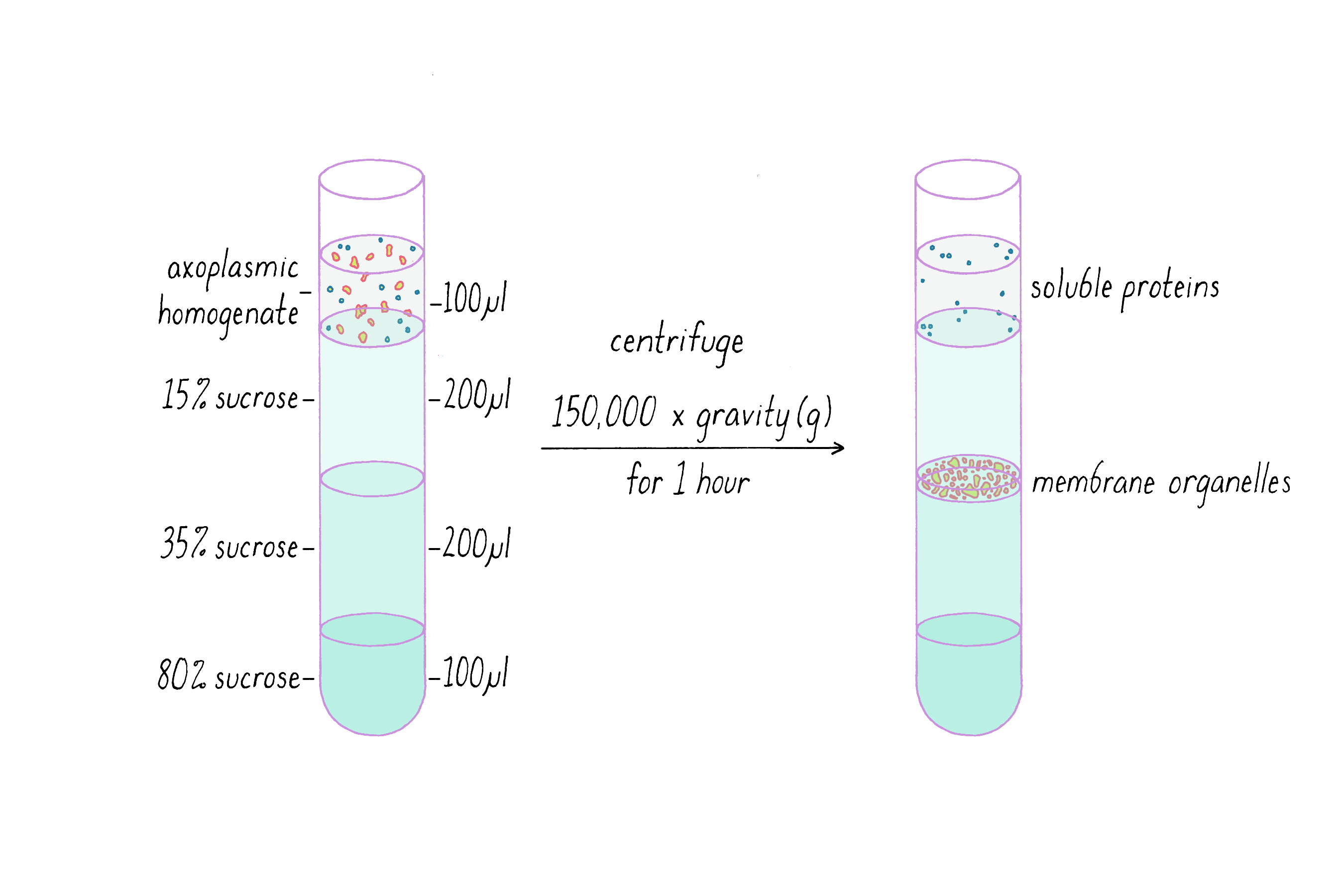
I was taught that a good biochemist initially keeps all fractions, because you may need to analyze them (and after all, you can always throw them away later). Thus, I also kept the soluble protein fraction that remained at the very top of the tube after centrifugation. This is a collection of thousands of different kinds of proteins that are not associated with organelles or cytoskeletal filaments. These proteins are small compared to organelles and less affected by the forces generated by ultracentrifugation (Figure 7). While I initially did not foresee their importance, the soluble proteins would end up playing a critical role in the experiment described below.
Doing the Key Experiment
Preparing the experimental materials described above was easier said than done. Some days, the collecting boat failed to return with squid. Other days, the microtubule preparation worked but the organelle preparation failed or vice versa. The axoplasm preparations also could be finicky and sometimes would lose organelle transport activity after being removed from the axon. Science requires a lot of persistence. Experimental failures are not necessarily "your failures"; trial and error is an inevitable part of science. However, fortunes can change from one day to the next, as revealed in this story (Video 7).
The First Experimental Attempt
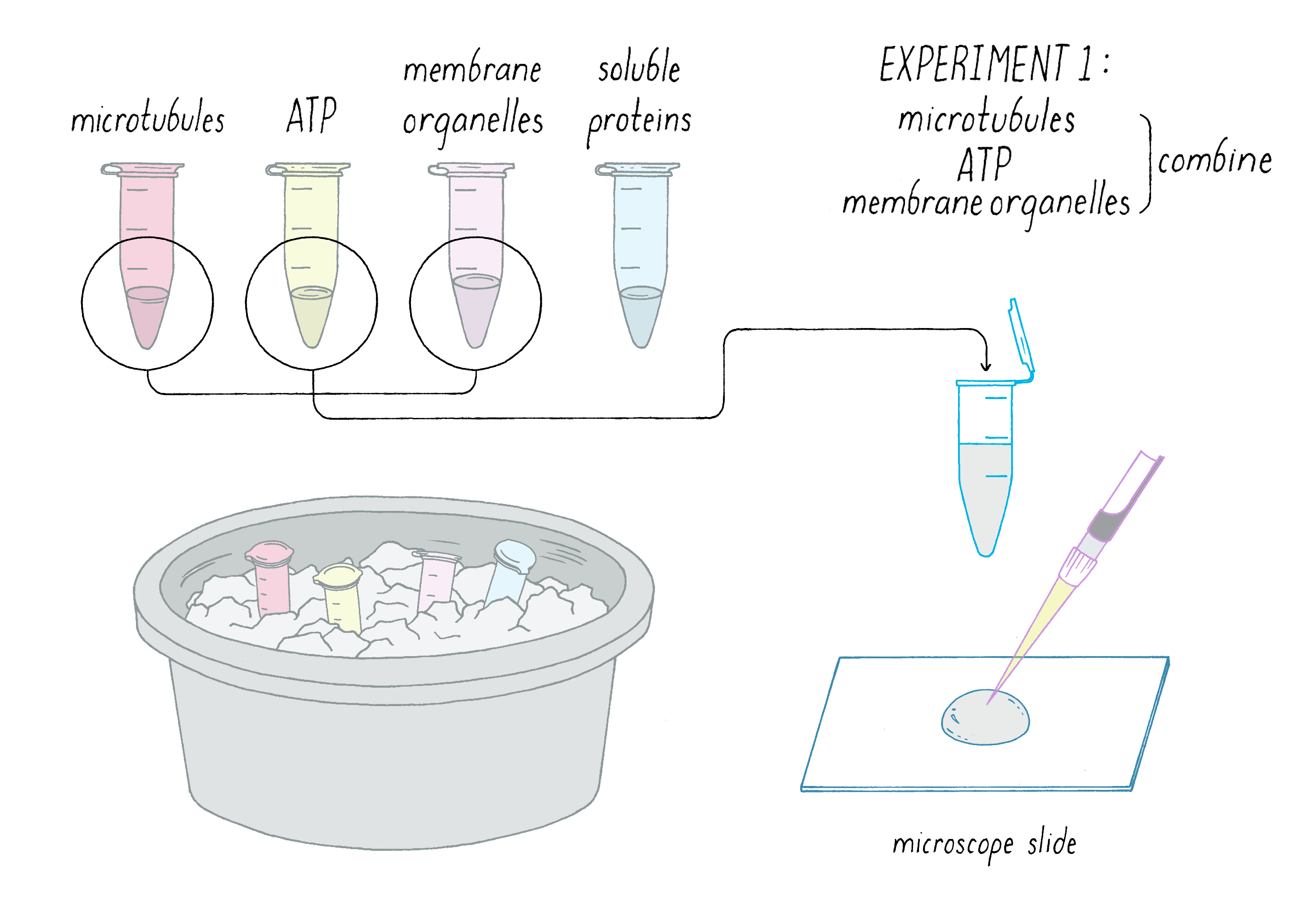
Finally, one day in the late summer of 1984, the preparation seemed to go well. After dinner, I had an ice bucket (biological samples retain their activity for longer when kept cold) with separate tubes of the materials. I mixed together what I thought were the key components for motility (Figure 8):
1) Microtubules
2) ATP
3) Membrane organelles
The mixture was added to a microscope slide, and I focused the video microscope with great anticipation. Would the axonal transport machinery come back to life? Would I see membrane organelles moving along microtubules?
Disappointing! Nothing happened. After months of work to get to this point, axonal transport in a test tube remained elusive.
The Second Experimental Attempt
When an experiment does not work, you have to think "why"? One possibility (true for most experiments) is purely technical—perhaps the method of preparation for one (or several) of the components was not good. The motor could have lost its activity after being outside of the cell for many hours.
Another possibility was that there was a missing ingredient. If so, might that missing ingredient be lurking in the soluble protein fraction? I had saved that fraction, and it was sitting in my ice bucket. Rather than admit defeat yet, why not try adding that?
So in Experiment 2 (Figure 9), I mixed together:
1) Microtubules
2) ATP
3) Membrane organelles
4) Soluble protein fraction

I was very excited by what I saw under the microscope: now the organelles were moving along the microtubules (similar to Video 3)!
This result suggested that something in the soluble protein fraction was necessary for the organelle movement. Before jumping to this conclusion, it was important to think of and rule out other possibilities. One nagging concern was that the moving objects were not membrane organelles but some other visible particle present in the soluble protein fraction. I had never looked at the soluble protein fraction through a microscope and thus could not rule out this possibility. It was time to perform a "control experiment." Let us take a moment to learn about control experiments, as they are the foundation of experimental biology. Controls test the contributions of different variables to an experimental outcome.
What are variables? Variables are essentially all of the specific components and conditions that make up an experiment. In the experiment discussed here, each component (microtubules, soluble protein, ATP, membrane organelles) is an experimental variable. Each of these components could be added or not; if added, their concentration could be adjusted. The components of the buffer solution (each chemical and its concentration, the pH, etc) also are variables. The temperature at which the experiment was performed is also a variable (temperature can affect the rates of reactions). As you can see, a single experiment can have many variables! A good experimental biologist will test what they think are likely to be important variables through positive and negative controls.
A positive control usually tests whether a certain variable(s) is behaving as expected. For example, what if I never saw organelles moving? Maybe I bought a bad batch of ATP?
Explorer's Question: How would you design a positive control to test your ATP?
Answer: There are various ways in which this could be done. You need a well-established and reliable test that requires ATP. One could test the ATP solution on a well-established ATPase enzyme. Or one could have the ATP chemically analyzed.
The negative control usually eliminates a variable to test whether it contributes to the experimental outcome.
Explorer's Question: In Experiment 2 (Figure 9), you postulate that the membrane organelle motility observed is powered by ATP. What negative control would test that hypothesis?
Answer: Make the mixture again but leave out ATP (so microtubules, organelles, soluble protein fraction only). If organelle motility is not observed (repeating the experiment several times), then you can conclude that ATP is required for motility. If motility continued, however, you could not necessarily immediately conclude that motility does not require ATP. An alternative explanation is that some ATP from the cell was present in the soluble protein fraction. You would have to do another experiment in which you tested for the presence of ATP in the soluble protein fraction or remove it. Experiments can be complicated and you need to think through many possibilities.
The Third Experimental Attempt
Back to our experiment, we postulated that the objects moving along microtubules were the membrane organelles, stimulated by a factor in the soluble protein fraction. How could we be sure that they were membrane organelles? Here the negative control was to eliminate the variable of the membrane organelles from the mixture.
In Experiment 3, I combined (Figure 10):
1) microtubules,
2) ATP, and
3) soluble protein fraction
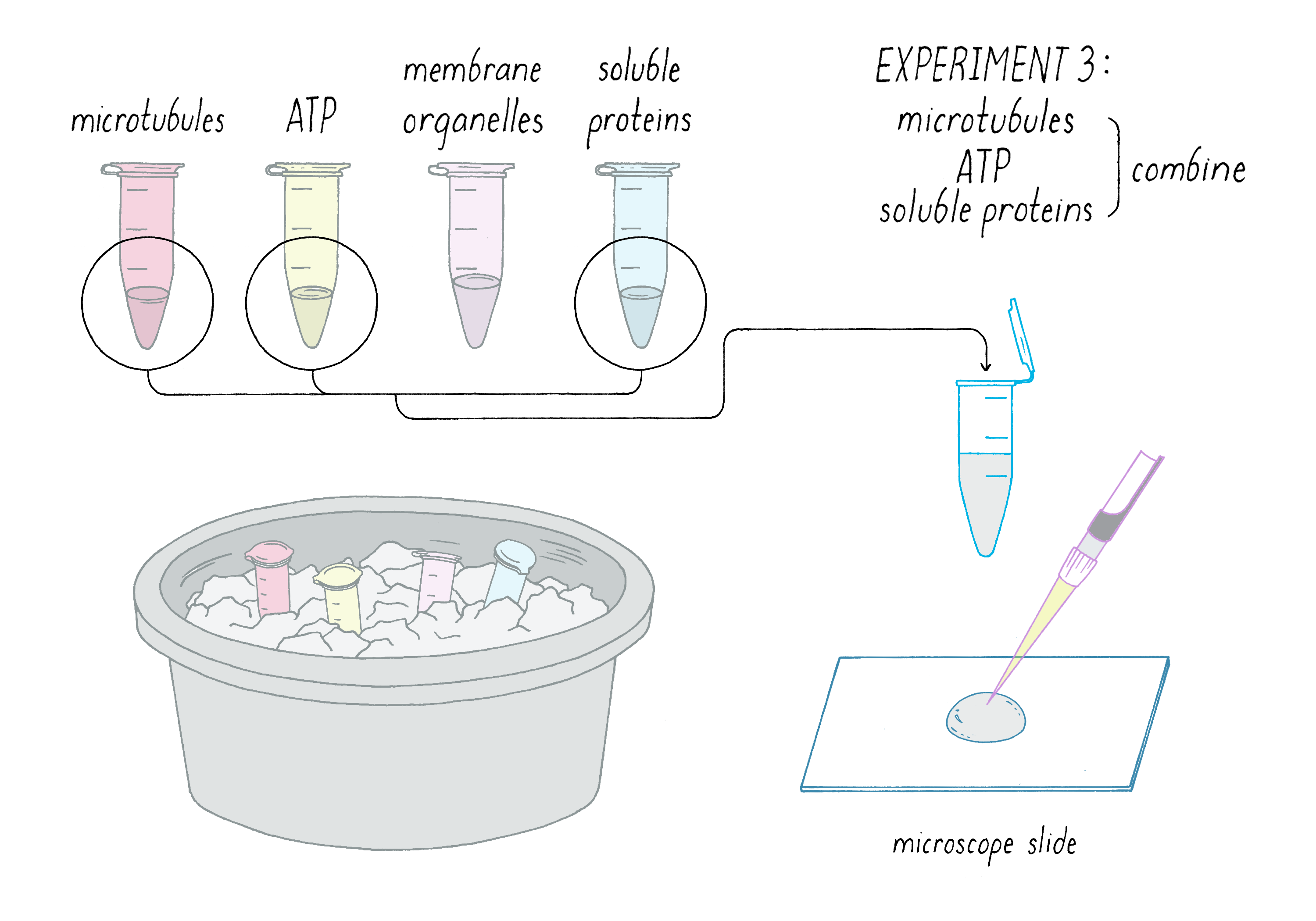
The expectation from this negative control was that I would see microtubules but nothing moving along them. Not very exciting, but I could go home at 1 AM feeling that a key control experiment was performed.
I looked under the microscope.
The video camera revealed microtubules crawling along the glass surface like worms (Video 8). This observation was mesmerizing but definitely not in the game plan. We expected organelles to move along stationary microtubule tracks. Clearly, this post-midnight control experiment revealed something unexpected. Perhaps it was opening a new door to the secret of axonal transport (see Dig Deeper 3 for additional comments on this experiment).
Lots of new questions arose in those first minutes of watching microtubules crawl on glass. Why were the microtubules moving without organelles?
Thinking quickly, a new hypothesis arose:
1) the soluble fraction contained motor proteins,
2) the motor proteins "stuck" onto the glass surface, and
3) motor proteins grabbed hold of the microtubule and pushed it in one direction.
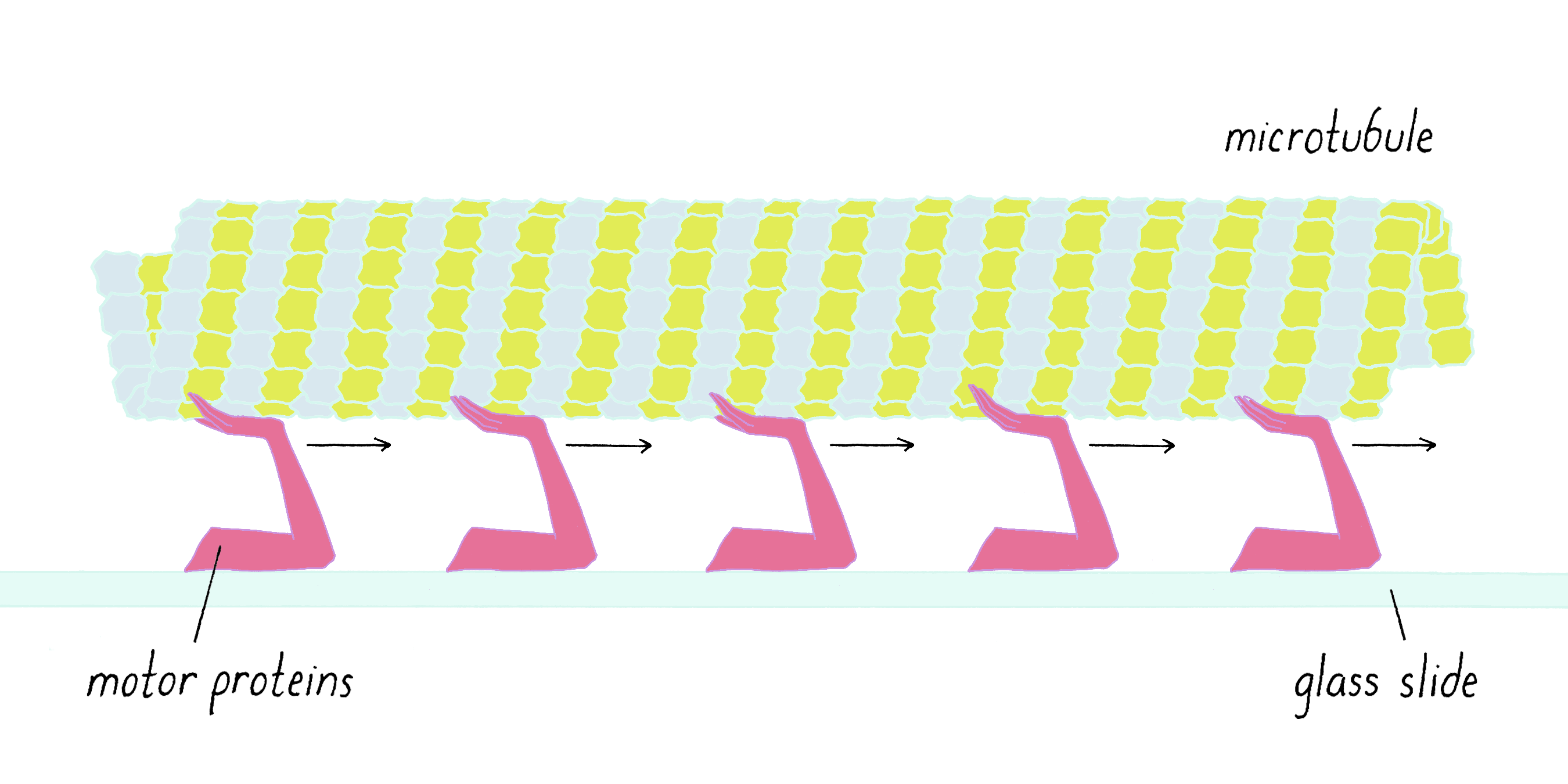
Because the motors were stuck onto the glass and could not move, the microtubules were moving relative to the stationary motors. Think "crowd surfing." The rock star is transported by people who are largely stationary in the mosh pit (Figure 11).

Perhaps the microtubules were moving in an analogous manner, with some moving arm from the stationary motor protein transporting the microtubule (Figure 12).
Additional Confirmatory Experiments
Sometimes a hypothesis might require years to test. However, the above hypothesis could be tested with a 15-minute experiment (Figure 13).
1) A drop of the soluble protein fraction was placed on the microscope slide for 10 min. Using a marker pen, the position of the drop of soluble proteins was labeled (A). A nearby area that was not exposed to the proteins was also labeled (B).
2) The slide was washed to remove all proteins that did not stick to the glass.
3) A solution of just microtubules and ATP was added to the entire slide, covering both areas A and B. The slide was examined by microscopy to see if microtubules were moving across the surface.

The result was clear (Figure 13). Microtubules were moving along the surface of glass in area A, where the soluble protein drop had been placed before being washed. The microtubule did not move in area B, which never was exposed to the soluble proteins. We concluded that some type of motor protein in the soluble protein fraction had attached firmly to the surface so that it was not washed away and remained active (Figure 13).
In the following week, we tested whether the mystery motor would stick to other surfaces. Amazingly, the motor also stuck to and transported very small plastic beads that were 1 µm in diameter (one-millionth of a meter). The motion looked very much like the transport of membrane organelles, the natural biological cargo (Video 9).
Explorer's Question: What features of the plastic bead motion do you notice in this movie?
Answer: There are two types of motion. One is straight and directed along the microtubule. This motion is driven by motor proteins. The other is random, "bouncy," fast motion in solution (also in 3D because the beads are coming in and out of focus). This is "Brownian Motion" which is driven by thermal energy of water molecules colliding with the beads. Brownian motion is discussed by Prakash in his Narrative. All the beads on the microtubule move in one direction. You might recall that there was bidirectional motion in Videos 1 and 3. The answer became evident in a later study. The motion shown here is caused by kinesin, which only moves in one direction. There is another motor that moves in the other direction, which was found to be a dynein. You may observe other interesting things as well in this movie.
After months of nothing working in the lab, now within a few days, lots of things were moving in a test tube environment. I had what I came for—an in vitro reconstitution of membrane organelles moving along purified microtubules. But two additional interesting and unexpected results had emerged: 1) microtubules being transported along glass by surface-bound motor proteins and 2) small plastic beads, with bound motor proteins, being transported along a stationary microtubule. We now had lots of "assays" for detecting biological motility. An assay is an experimental method for measuring a very specific biological activity. In our case, serendipity had pointed us to new assays to measure the motion produced by an invisible motor protein. Assays are powerful tools for researchers, and you will encounter them frequently in experimental biology.
In the next section, I will explain why these assays proved to be a scientific goldmine.
What happened next?
After these amazing few days of experiments, I called Stanford and asked if I could postpone my return to medical school. I wanted to hunt for the motor that was moving beads along microtubules and moving microtubules across the microscope slide. Presumably, the same motor was transporting organelles as well. Stanford granted my request.
Using the new in vitro motility assays described above, I purified a new molecular motor protein a few months later and called it "kinesin". In the subsequent decade, work from many scientists revealed that kinesin is not a single motor but a family of related motor proteins (Video 10). Humans have 45 different kinesins that are involved in many aspects of biology, including development, cell division, and cell organization, in addition to axonal transport. A couple of years later, Richard Vallee found a dynein motor in axons (a version of a motor protein that was first discovered in cilia and flagella) using the in vitro motility assays described here.
The assays described here became "work horses" for discovering new motor proteins and understanding how motor proteins work. Compared to studying motility in the complicated environment of the intact cells with thousands of proteins, the simplicity of this system was powerful. All that is needed is a purified motor protein, a purified microtubule, and an artificial substrate such as a bead or glass surface (see Dig Deeper 4 for why we were incredibly lucky that these artificial substrates worked in activating the kinesin motor protein). These assays also joined two fields together—microscopy (previously used for looking at cells) with biochemistry (the study of molecules). Now direct observation by microscopy could be used to understand how proteins work.
There were many new exciting problems to solve with these approaches. How many motor proteins were required to move microtubules? To answer this question, the number of motors bound to the glass was progressively decreased to a point where there was only a single kinesin protein in contact with the microtubule. Amazingly, a single kinesin could happily transport a microtubule that was 1000 times longer than itself. Thus, the assays were not only simple but also extraordinarily sensitive; they could detect motion from a single motor protein. An entire field of single molecule biophysics was born. By studying individual motor proteins, several laboratories found that motors take discrete steps along a track for each ATP molecule consumed.
So what happened next in my career? I never did return to medical school; there was never a convenient time to stop my research. There were always more questions that stimulated my curiosity; it has turned out to be a non-stop science adventure ride ever since. Instead of returning to medical school, I took a job as an assistant professor at University of California, San Francisco. Just like science, the path of one's life can be unpredictable.
Closing Thoughts
The "scientific method" is often presented in textbooks as a flow chart beginning with a hypothesis followed by an experiment that proves that hypothesis right or wrong. This linear and logical path fails to capture the twists, turns, and surprises that can occur in research. Our experiment was designed to reconstitute the movement of organelles along microtubules, but the most exciting result, microtubules moving on glass surfaces, was unplanned and unexpected. The original hypothesis was not necessarily right or wrong; it was incomplete.
In this "Key Experiment," the unexpected result of microtubules moving on the glass was very obvious when we encountered it. We also knew immediately that it was important and how it could be exploited to move the project forward. However, on other occasions, I missed opportunities to recognize and capitalize on an unexpected result. On yet other occasions, I followed unexpected results into blind alleys. Recognizing clues and following them down a path toward a new discovery is not always easy. Two scientists can look at the same set of clues in very different ways and take different paths. This element of intuition makes science creative.
Why was this experiment a surprise? Why didn't we conceive of the "exciting experiment" from the beginning, instead of stumbling across it? Consider these two explanations.
First, scientists, being human, have biases as to what they expect from an experiment. A hypothesis is a form of a preconceived notion, which may be incorrect or, at minimum, missing elements of the full story. In our case, we thought that motors would be permanently bound to organelles; we did not consider that there would be a large pool of motors unattached to any cargo. In retrospect, having a reserve pool of motors held in an inactive state makes a lot of sense (see Dig Deeper 4), but we did not think of it at the time.
Second, it is very difficult to envision the unknown, until….. one bumps into it. Scientific research, by its nature, probes the murky interface between the known and the unknown. Like Christopher Columbus who "bumped into North America" on his intended route to the Indies, unexpected experimental results take scientists into new territories that they did not anticipate or even realize existed. Bumping into something new usually proves to be more interesting than the original intended journey. Such unpredictability in the search for new knowledge makes science exciting and deeply interesting.
Dig Deeper
Dig Deeper 1: Preparation of microtubules from purified tubulin
Biochemistry utilizes techniques of fractionation, which results in the purification of a single protein from an initial complex mixture of thousands. To obtain purified tubulin, we started with squid optic lobes (the squid brain), because they were easier to obtain in large quantities than the giant axon. The details can be found in the Guided Paper. Once we had the purified tubulin, we could use certain buffers (see Dig Deeper 2) that cause the tubulin to assemble into microtubules. Figure DD1A shows an electron microscopy image of microtubules after they have been assembled in a test tube. Each of these microtubules are ~10–20 micrometers in length and contain >16,000 copies of the tubulin protein. Microtubules also can depolymerize back to tubulin. To prevent the microtubule structure from falling apart while measuring organelle transport, we used a microtubule-stabilizing drug called taxol, which is also used in cancer treatment.

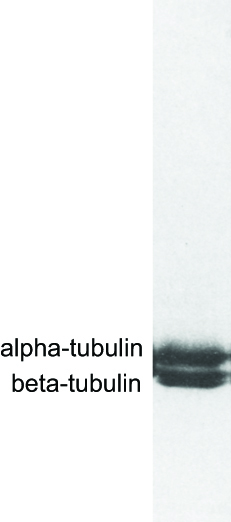
How can one assess the purity of the tubulin made from squid optic lobes? One can perform a technique called polyacrylamide gel electrophoresis, which separates proteins based upon their size. The method is analogous to separation of different-sized DNA molecules by agarose gel electrophoresis, which is described in this Video and the interactive Activity by Shawn Douglas. The separated proteins are visualized with a protein-binding dye (Figure DD1B).
In the figure below, two bands, corresponding to alpha-tubulin and beta-tubulin, are seen but little else. More sensitive techniques would reveal minor other contaminating proteins, but for the purposes of this experiment, the tubulin preparation was considered to be relatively "clean" and devoid of microtubule-associated proteins.
Dig Deeper 2: Buffers
Biochemists prepare very specific solutions for their in vitro reactions. Such a solution is commonly referred to as a "buffer" or "buffer solution." A buffer, in its more strict chemical definition from Wikipedia, is "a solution that resists changes in pH when acid or alkali is added to it. Buffers typically involve a weak acid or alkali together with one of its salts." Setting the pH, and preventing fluctuations of pH, is an important aspect of biochemical buffers. The pH (hydrogen ion concentration) is usually adjusted to a value in the physiological range (between 6 and 8). The buffering agents used in these experiments included small organic molecules that were previously identified in squid axons as well as a human-made buffering agent (HEPES); the overall pH was adjusted to 7.2.
In addition to the agent used to set and buffer the pH, buffer solutions contain salts composed of positively and negatively charged ions. Common positive cations are potassium (K+; the major cation inside of cells) or sodium (Na+; the major cation in human blood or sea water). Common negative anions include chloride (Cl−), acetate (C2H3O−) or other organic components. Buffers may include divalent cations such as magnesium (Mg2+). Magnesium is essential for many enzymes in the cell, particularly ATPases such as the motor proteins featured in this experiment. The calcium concentration in cells is very low, so sometimes calcium buffers (EGTA) are used to absorb or maintain calcium at low levels.
A buffer is made by weighing out chemicals, dissolving them in pure water to a defined concentration, and then adjusting the pH with acid or base. Establishing the right buffer conditions is sometimes done by varying one component systematically and then examining the effect on the in vitro biochemical activity. Finding the right buffer conditions is not trivial. Like a great chef who picks the right ingredients for a dish, buffers are part of the "art" of biochemistry. Choosing the right buffer ingredients and their concentrations can make the difference between success and failure of a scientific project. Indeed, finding the right buffer conditions was critical for the in vitro reconstitution of organelle transport. Much of this work was established in a publication that preceded this "Key Experiment."
Dig Deeper 3: Additional insights on the experiments
You might be curious about the following:
Why did we not see microtubules moving on the glass in Experiment 2 when the organelles were also present?
Actually, they were moving in that experiment too (as we found out later), but we were distracted by the movement of membrane organelles being transported along the microtubules, which was the phenomenon that we were looking for. There is a famous psychology video on how one can miss an unexpected phenomenon by focusing on an expected event. https://www.youtube.com/watch?v=IGQmdoK_ZfY
If the motors are randomly distributed on the glass (as one would expect), why do the microtubule move unidirectionally? Why don't the randomly placed motors pull the microtubules in opposite directions and the tug-of-war causing them to a halt?
Microtubules are polar structures, with a distinct "plus" and "minus" end. This is because tubulin proteins (which themselves are asymmetric) are all aligned in a similar head-to-tail manner. The motors recognize the microtubule polarity and will only attach in one orientation. Thus, motors facing the "wrong direction" on the microtubule will not attach (unless they swivel around by 180°, which they might do). Thus, the motors oriented in the direction along the microtubule path will all produce movement in the same direction. The ones in the opposite direction will not produce any action on the microtubule.
The velocity of microtubules moving on glass looks slower than the velocity of organelles moving on the microtubules (also reported in the Guided Paper). Why is this?
This still remains an interesting and unanswered question! We now know that there are many kinesin motors in an axon not just one. "Conventional kinesin" (Kinesin 1) is the dominant motor in the soluble protein fraction and the one primarily responsible for moving microtubules on glass. But there is another motor called Kinesin 3, which is threefold to fourfold faster than Kinesin 1. Many of the small, fast moving membrane organelles might be transported by Kinesin 3 while the slower ones (e.g., mitochondria) might be moved by Kinesin 1. However, there are also reports that Kinesin 1-driven transport can be faster than its transport of microtubules on glass. Perhaps there is an activation mechanism that accelerates its speed? There are still many open questions to solve.
Dig Deeper 4: Motor protein regulation and the good fortune of our experiment
The unexpected in vitro motility results raised a puzzling question: why were there so many molecular motors floating around free in the cytoplasm and not attached to any membrane organelle cargo? It seemed wasteful to have motors burning chemical energy (ATP hydrolysis) without producing movement.
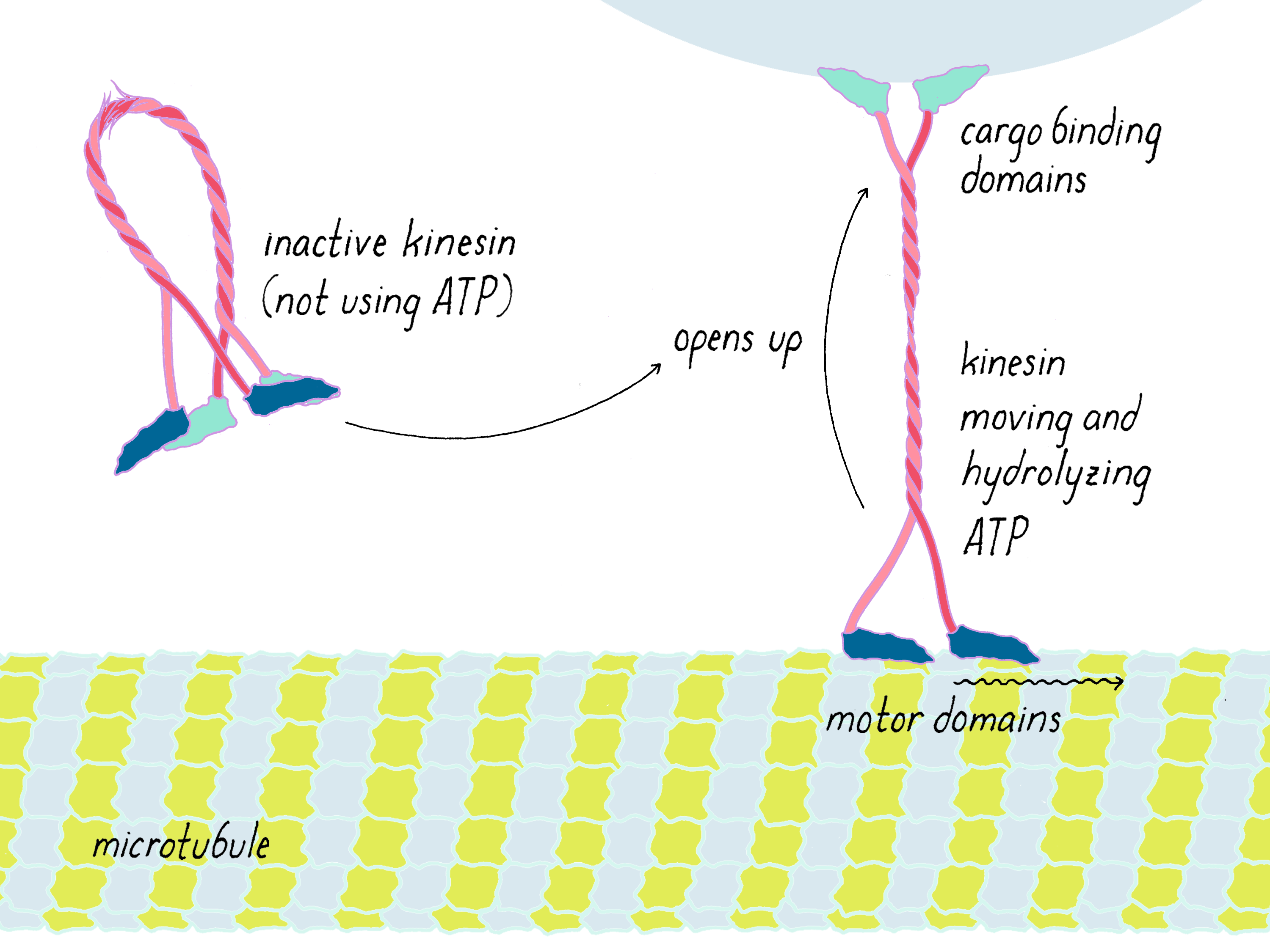
It was later discovered that nature is indeed not wasteful; rather we were just lucky in our experiment. Normally, motor proteins are turned off (not burning ATP) when they are unattached to cargo in the soluble protein fraction and then turned on when they bind to a membrane organelle cargo (Figure DD4). Kinesin is a long molecule with two different ends; one end contains the motor domains that walk along the microtubule and the other end attaches to proteins on an organelle. When detached from its cargo, kinesin folds in half like a gymnast through a flexible joint in the middle; the organelle binding domain grabs hold of the motor domain and prevents it from hydrolyzing ATP. Binding to the membrane organelle "unfolds" kinesin, so that it can now start consuming ATP and carry its cargo along the microtubule.
So why did we see movement in our experiments if kinesin motors in the soluble protein fraction are turned off? We think, although it is not proven, that negative charges on the glass surface of the beads interact with positive-charged amino acids in the cargo binding domain; in doing so, the motor unfolds from its inactive conformation and is switched on.
The bottom line: we were incredibly lucky that artificial surfaces activated the motor. If the motors were turned off in the soluble protein fraction, we would have never been able to detect them. The pivotal control experiment described in this "Key Experiment" would have yielded no surprises. I would have still reconstituted organelle transport, but the other assays would not have been discovered. Without them, the next steps to identify the motor would have been hard and non-obvious. Recognizing that, I likely would have returned to medical school. Nature normally does not give away its secrets easily but occasionally one gets a lucky break.
References and Resources
Guided Paper
Vale, R.D., Schnapp, B.J, Reese, T.S. and Sheetz, M.P. Organelle, bead and microtubule translocations promoted by soluble factors from the squid giant axon. Cell 1985;40:559–569.
This paper shows that a protein, now called kinesin, is responsible for the complex process of membrane organelle transport within nerve cells, providing new insights and tools for understanding the basis of biological motility.
DownloadResources
- An iBiology talk on the discovery of kinesin by Ron Vale. https://www.ibiology.org/cell-biology/kinesin/
This video covers similar material to this Key Experiment.
- An iBiology talk by Ron Vale on Molecular Motor Proteins. Available at: https://www.ibiology.org/cell-biology/motor-proteins/
This video, by Ron Vale, provides an overview into the biology of motor proteins.
- What in Kinesin? (in 3.5 min by Ron Vale). Available at: https://www.youtube.com/watch?v=mBo_o0iO68U
- The Life of a Motor Protein. Available at: https://www.youtube.com/watch?v=tMKlPDBRJ1E
This short and fun video describes how organelles filled with cellular materials are transported throughout a nerve cell via motor proteins.
- Differential interference contrast microscopy (the technique used to image the microtubules and membrane organelles); iBiology talk by Ted Salmon. Available at: https://www.ibiology.org/talks/differential-interference-contrast/
- Explanation of the Buoyancy Force by the Khan Academy. Available at: https://www.khanacademy.org/science/physics/fluids/buoyant-force-and-archimedes-principle/v/fluids-part-5
- Concept of centripetal and centrifugal forces (relevant to ultracentrifugaton) by the Khan Academy. Available at: https://www.khanacademy.org/science/ap-physics-1/ap-centripetal-force-and-gravitation/centripetal-forces-ap/v/introduction-to-centripetal-force
- Animated Videos of Kinesin Walking Along Microtubules—Inner Life of the Cell, made by BioVisions at Harvard University, features the kinesin motor protein. Available at: https://www.youtube.com/watch?v=wJyUtbn0O5Y
- The Workhorse of the Cell: Kinesin. Available at: https://www.youtube.com/watch?v=gbycQf1TbM0.
- Narrated version of Kinesin walking and conversion of ATP energy into motion. Available at: https://www.youtube.com/watch?v=YAva4g3Pk6k
- Transport inside the brain—The basic mechanisms of neuronal trafficking. Available at: https://www.youtube.com/watch?v=RRfH4ixgJwg